
Visi iLive saturs ir medicīniski pārskatīts vai pārbaudīts, lai nodrošinātu pēc iespējas lielāku faktisko precizitāti.
Mums ir stingras iegādes vadlīnijas un tikai saikne ar cienījamiem mediju portāliem, akadēmiskām pētniecības iestādēm un, ja vien iespējams, medicīniski salīdzinošiem pārskatiem. Ņemiet vērā, ka iekavās ([1], [2] uc) esošie numuri ir klikšķi uz šīm studijām.
Ja uzskatāt, ka kāds no mūsu saturiem ir neprecīzs, novecojis vai citādi apšaubāms, lūdzu, atlasiet to un nospiediet Ctrl + Enter.
Hipotalāmu
Raksta medicīnas eksperts

Pēdējā pārskatīšana: 07.07.2025
Hipotalāms veido diencefalona apakšējās daļas un piedalās trešā kambara pamatnes veidošanā. Hipotalāmā ietilpst redzes nerva hiasma, redzes trakts, pelēkais bumbulis ar piltuvi un piena dziedzeru ķermeņi.
Redzes nerva hiazma (chiasma opticum) ir šķērsvirziena izvirzījums, ko veido redzes nervu (II galvaskausa nervu pāris) šķiedras, daļēji šķērsojot tās uz pretējo pusi (veidojot dekusāciju). Šis izvirzījums turpinās laterāli un aizmugures virzienā katrā pusē redzes traktā (tratus opticus). Redzes trakts atrodas mediāli un aizmugures virzienā no priekšējās perforētās vielas, no laterālās puses liecas ap smadzeņu kājiņu un beidzas ar divām saknēm subkortikālajos redzes centros. Lielākā laterālā sakne (radix lateralis) tuvojas laterālajam ģenikulārajam ķermenim, bet plānākā mediālā sakne (radix medialis) iet uz vidussmadzeņu jumta augšējo colliculus kaulu.
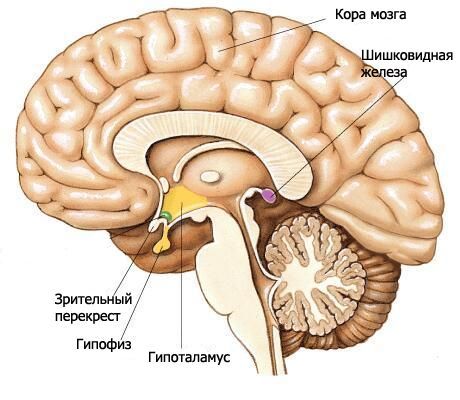
Termināla plāksne, kas pieder telencephalonam, atrodas blakus redzes nerva hiasmas priekšējai virsmai un saplūst ar to. Tā noslēdz smadzeņu gareniskās spraugas priekšējo daļu un sastāv no plāna pelēkās vielas slāņa, kas plāksnes sānu daļās turpinās pusložu frontālās daivas vielā.
Aiz redzes nerva hiazmas atrodas pelēkais bumbulis (tuber cinereum), aiz kura atrodas piena dziedzeru ķermeņi, bet sānos - redzes trakti. Zemāk pelēkais bumbulis pāriet piltuvē (infundibulum), kas savienojas ar hipofīzi. Pelēkā bumbuļa sienas veido plāna pelēkās vielas plāksnīte, kurā atrodas pelēkie bumbuļveida kodoli (nuclei tuberales). No trešā kambara dobuma puses sašaurinoša piltuves ieplaka izvirzīta pelēkā bumbuļa rajonā un tālāk piltuvē.
Zīdkoka ķermeņi (corpora mamillaria) atrodas starp pelēko bumbuli priekšpusē un aizmugurējo perforēto vielu aiz muguras. Tie izskatās kā divi mazi, apmēram 0,5 cm diametrā, sfēriski balti veidojumi. Baltā viela atrodas tikai ārpus zīdkoka ķermeņa. Iekšpusē atrodas pelēkā viela, kurā izšķir zīdkoka ķermeņa mediālo un laterālo kodolu (nuclei corporis mamillaris mediales et laterales). Forniksa kolonnas beidzas zīdkoka ķermeņos.
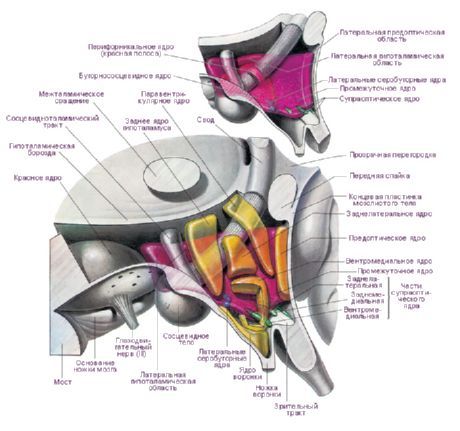
Hipotalāmā ir trīs galvenie hipotalāma reģioni - dažādu formu un izmēru nervu šūnu grupu kopas: priekšējais (regio hypothalamica anterior), starpposma (regio hypothalamica intermedia) un aizmugurējais (regio hypothalamica posterior). Nervu šūnu kopas šajos reģionos veido vairāk nekā 30 hipotalāma kodolus.
Hipotalāma kodolu nervu šūnām piemīt spēja producēt sekrētu (neirosekrēciju), ko caur šo pašu šūnu izaugumiem var transportēt uz hipofīzi. Šādus kodolus sauc par hipotalāma neirosekrēcijas kodoliem. Hipotalāma priekšējā rajonā atrodas supraoptiskais (supraoptiskais) kodols (nucleus supraopticus) un paraventrikulārie kodoli (nuclei paraventriculares). Šo kodolu šūnu izaugumi veido hipotalāma-hipofīzes saišķi, kas beidzas hipofīzes aizmugurējā daivā. Starp hipotalāma aizmugurējā rajona kodolu grupu lielākie ir piena dziedzera ķermeņa mediālais un laterālais kodols (nuclei corporis mamillaris mediales et laterales) un hipotalāma aizmugurējais kodols (nucleus hypothalamicus posterior). Starpposma hipotalāma reģiona kodolu grupā ietilpst inferomediālie un superomediālie hipotalāma kodoli (nuclei hypothalamic ventromediales et dorsomediales), dorsālais hipotalāma kodols (nucleus hypothalamicus dorsalis), infundibulara kodols (nucleus infundibularis), pelēkbumbveida kodoli (nuclei tuberales) utt.
Hipotalāmu kodolus savieno diezgan sarežģīta aferentu un eferentu ceļu sistēma. Tādēļ hipotalāmam ir regulējoša ietekme uz daudzām organisma veģetatīvajām funkcijām. Hipotalāmu kodolu neirosekrēcija var ietekmēt hipofīzes dziedzeru šūnu funkcijas, palielinot vai kavējot vairāku hormonu sekrēciju, kas savukārt regulē citu endokrīno dziedzeru aktivitāti.
Neironu un humorālo savienojumu klātbūtne starp hipotalāma kodoliem un hipofīzi ļāva tos apvienot hipotalāma-hipofīzes sistēmā.
Filoģenētiskie pētījumi ir parādījuši, ka hipotalāms pastāv visiem hordaiņiem, ir labi attīstīts abiniekiem un vēl jo vairāk rāpuļiem un zivīm. Putniem ir skaidri izteikta kodolu diferenciācija. Zīdītājiem pelēkā viela ir ļoti attīstīta, kuras šūnas diferencējas kodolos un laukos. Cilvēka hipotalāms būtiski neatšķiras no augstāko zīdītāju hipotalāma.
Pastāv liels skaits hipotalāma kodolu klasifikāciju. E. Gruntels identificēja 15 kodolu pārus, W. Le Gros Clark - 16, H. Kulenbeks - 29. Visplašāk izmantotā klasifikācija ir W. Le Gros Clark klasifikācija. I. N. Bogolepova (1968), pamatojoties uz iepriekš minētajām klasifikācijām un ņemot vērā ontoģenēzes datus, ierosina sadalīt hipotalāma kodolus četrās daļās:
- priekšējā jeb rostrālā daļa (apvienojot preoptisko zonu un priekšējo grupu - W. Le Gros Clark) - preoptiskās mediālās un sānu zonas, suprachiasmatiskais kodols, supraoptiskais kodols, paraventrikulārais kodols, priekšējā hipotalāma zona;
- vidējā mediālā daļa - ventromediālais kodols, dorsomediālais kodols, infundibulārais kodols, aizmugurējā hipotalāma zona;
- vidējā sānu daļa - laterālā hipotalāma zona, laterālais hipotalāma kodols, tuberolaterālais kodols, tuberomamimilārais kodols, periforniskais kodols;
- aizmugurējā jeb mamilārā daļa - mediālais mamilārais kodols, sānu mamilārais kodols.
Hipotalāma anatomiskie savienojumi arī paskaidro tā (funkcionālo) nozīmi. Starp svarīgākajiem aferentajiem ceļiem var izdalīt šādus:
- mediālais priekšsmadzeņu saišķis, kura laterālā daļa savieno hipotalāmu ar ožas sīpolu un bumbuli, periamigdaloīdu reģionu un hipokampu, bet mediālā daļa — ar smadzeņu starpsienu, diagonālo reģionu un astes kodolu;
- termināla sloksne, kas stiepjas no mandeles līdz hipotalāma priekšējām daļām;
- šķiedras, kas iet caur fornix no hipokampa līdz piena dziedzeru ķermenim;
- talamo-, strio- un pallidohipotalāma savienojumi;
- no smadzeņu stumbra - centrālais tegmentālais trakts;
- no smadzeņu garozas (orbitālā, temporālā, parietālā).
Tādējādi galvenie aferentācijas avoti ir priekšsmadzeņu limbiskie veidojumi un smadzeņu stumbra retikulārais veidojums.
Hipotalāmu eferentās sistēmas var iedalīt arī trīs virzienos:
- dilstošās sistēmas uz retikulāro formāciju un muguras smadzenēm - periventrikulārā šķiedru sistēma, kas beidzas vidussmadzenēs (gareniskais aizmugurējais saišķis), astes stumbra un muguras smadzeņu autonomajos centros, un piena dziedzeru-tegmentālais saišķis, kas iet no piena dziedzeru ķermeņiem uz vidussmadzeņu retikulāro formāciju;
- ceļi uz talāmu no piena dziedzeru ķermeņiem (mammatalāma saišķis), kas ir daļa no slēgtās funkcionālās limbiskās sistēmas;
- ceļi uz hipofīzi - hipotalāma-hipofīzes ceļš no paraventrikulārajiem (10-20% šķiedru) un supraoptiskajiem (80-90%) kodoliem līdz hipofīzes aizmugurējām un daļēji vidējām daivām, tuberohipofīzes ceļš no ventromediālajiem un infundibulārajiem kodoliem līdz adenohipofīzei.
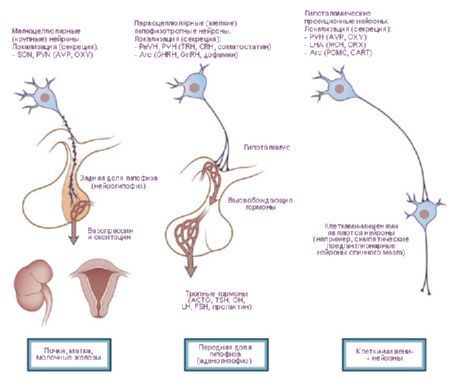
Dž. Ransona (1935) un V. Hesa (1930, 1954, 1968) darbos tika sniegti dati par zīlītes paplašināšanos un sašaurināšanos, arteriālā spiediena palielināšanos un samazināšanos, pulsa paātrināšanos un palēnināšanos, stimulējot hipotalāmu. Pamatojoties uz šiem pētījumiem, tika identificētas zonas, kurām ir simpātiska (hipotalāma aizmugurējā daļa) un parasimpātiska (priekšējā daļa) iedarbība, un pats hipotalāms tika uzskatīts par centru, kas integrē orgānus un audus inervējošās viscerālās sistēmas aktivitāti. Tomēr, attīstoties šiem pētījumiem, tika atklāts arī liels skaits somatisko efektu, īpaši dzīvnieku brīvas uzvedības laikā [Gellhorn E., 1948]. O. G. Baklavadžans (1969), stimulējot dažādas hipotalāma daļas, dažos gadījumos novēroja aktivācijas reakciju smadzeņu garozā, muguras smadzeņu monosinaptisko potenciālu veicināšanos, arteriālā spiediena paaugstināšanos, bet citos - pretēju efektu. Šajā gadījumā visaugstākais slieksnis bija veģetatīvajām reakcijām. O. Seigers (1962) atklāja y-sistēmas un EEG sinhronizācijas inhibīciju hipotalāma diatermijas laikā un pretēju efektu pārmērīgas sasilšanas laikā. Veidojas ideja par hipotalāmu kā smadzeņu daļu, kas veic mijiedarbību starp regulēšanas mehānismiem, somatiskās un veģetatīvās aktivitātes integrāciju. No šī viedokļa ir pareizāk hipotalāmu sadalīt nevis simpātiskajā un parasimpātiskajā daļā, bet gan tajā atšķirt dinamogēnās (ergotropās un trofotropās) zonas. Šī klasifikācija ir funkcionāla, bioloģiska pēc savas būtības un atspoguļo hipotalāma līdzdalību holistisku uzvedības aktu īstenošanā. Acīmredzot homeostāzes uzturēšanā piedalās ne tikai veģetatīvā, bet arī somatiskā sistēma. Ergotropās un trofotropās zonas atrodas visās hipotalāma daļās un dažās zonās pārklājas. Tajā pašā laikā ir iespējams identificēt to "koncentrācijas" zonas. Tādējādi priekšējās daļās (preoptiskajā zonā) skaidrāk ir attēloti trofotropiskie aparāti, bet aizmugurējās daļās (mamilārajos ķermeņos) - ergotropiskie. Hipotalāma galveno aferento un eferento savienojumu analīze ar limbisko un retikulāro sistēmu atklāj tā lomu integratīvo uzvedības formu organizēšanā. Hipotalāms šajā sistēmā ieņem īpašu - centrālu - pozīciju gan tā topogrāfiskā novietojuma dēļ šo veidojumu centrā, gan fizioloģisko īpašību rezultātā. Pēdējo nosaka hipotalāma loma kā īpaši konstruētai smadzeņu daļai, kas ir īpaši jutīga pret izmaiņām ķermeņa iekšējā vidē, reaģējot uz mazākajām humorālo rādītāju svārstībām un veidojot lietderīgas uzvedības darbības, reaģējot uz šīm izmaiņām.Hipotalāma īpašo lomu nosaka tā anatomiskais un funkcionālais tuvums hipofīzei. Hipotalāma kodoli tiek iedalīti specifiskos un nespecifiskos. Pirmajā grupā ietilpst veidojumi, kas projicējas uz hipofīzi, pārējie ietver citus kodolus, kuru stimulācijas ietekme var atšķirties atkarībā no trieciena stipruma. Hipotalāma specifiskajiem kodoliem ir skaidra ietekme un tie atšķiras no citiem smadzeņu veidojumiem ar spēju veikt neirokrīniju. Tie ietver pelēkā bumbuļa supraoptisko, paraventrikulāro un parvocelulāro kodolu. Tika konstatēts, ka antidiurētiskais hormons (ADH) veidojas supraoptiskajā un paraventrikulārajā kodolā, kas pa hipotalāma-hipofīzes trakta aksoniem nolaižas uz hipofīzes aizmugurējo daivu. Vēlāk tika pierādīts, ka hipotalāma neironos veidojas atbrīvojošie faktori, kas, nonākot adenohipofīzē, regulē trīskāršo hormonu sekrēciju: adrenokortikotropo (AKTH), luteinizējošo (LH), folikulus stimulējošo (FSH) un vairogdziedzeri stimulējošo (TSH). AKTH un TSH ieviešanas faktoru veidošanās zonas ir vidējā paugura priekšējās daļas un preoptiskā apgabala kodoli, bet GTG - pelēkā bumbuļa aizmugurējās daļas. Ir konstatēts, ka cilvēka hipotalāma-hipofīzes saišķos ir aptuveni 1 miljons nervu šķiedru.
Neapšaubāmi, neiroendokrīnajā regulācijā piedalās arī citas smadzeņu daļas (temporālā reģiona mediāli bazālās struktūras, smadzeņu stumbra retikulārais veidojums). Tomēr specifiskākais aparāts ir hipotalāms, kas ietver endokrīnās dziedzerus organisma integrālo reakciju sistēmā, īpaši stresa rakstura reakcijās. Trofo- un ergotropo sistēmu rīcībā aktivitātes nodrošināšanai ir ne tikai perifērās simpātiskās un parasimpātiskās sistēmas, bet arī specifiski neirohormonālie aparāti. Hipotalāma-hipofīzes sistēma, kas darbojas pēc atgriezeniskās saites principa, lielā mērā ir pašregulējoša. Īstenojošo faktoru veidošanās aktivitāti nosaka arī hormonu līmenis perifērajās asinīs.
Tādējādi hipotalāms ir svarīga smadzeņu limbiskās un retikulārās sistēmas sastāvdaļa, taču, iekļaujoties šajās sistēmās, tā saglabā savas specifiskās “ieejas” īpašas jutības veidā pret izmaiņām iekšējā vidē, kā arī specifiskas “izejas” caur hipotalāma-hipofīzes sistēmu, paraventrikulāriem savienojumiem ar zemāk esošajiem veģetatīvajiem veidojumiem, kā arī caur smadzeņu stumbra talāmu un retikulāro veidojumu uz garozu un muguras smadzenēm.
 [
[