
Visi iLive saturs ir medicīniski pārskatīts vai pārbaudīts, lai nodrošinātu pēc iespējas lielāku faktisko precizitāti.
Mums ir stingras iegādes vadlīnijas un tikai saikne ar cienījamiem mediju portāliem, akadēmiskām pētniecības iestādēm un, ja vien iespējams, medicīniski salīdzinošiem pārskatiem. Ņemiet vērā, ka iekavās ([1], [2] uc) esošie numuri ir klikšķi uz šīm studijām.
Ja uzskatāt, ka kāds no mūsu saturiem ir neprecīzs, novecojis vai citādi apšaubāms, lūdzu, atlasiet to un nospiediet Ctrl + Enter.
Olnīcas
Raksta medicīnas eksperts

Pēdējā pārskatīšana: 04.07.2025

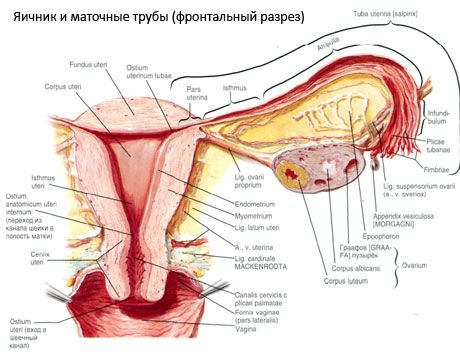
Olnīca (ovarium; grieķu val. oophoron) ir pāru orgāns, sieviešu reproduktīvais dziedzeris, kas atrodas iegurņa dobumā aiz dzemdes platās saites. Olnīcās attīstās un nobriest sieviešu reproduktīvās šūnas (olšūnas), un veidojas sieviešu dzimumhormoni, kas nonāk asinīs un limfā. Olnīcai ir olveida forma, nedaudz saplacināta anteroposterior virzienā. Olnīcas krāsa ir rozīga. Sievietei, kas dzemdējusi, olnīcas virsmā ir redzamas ieplakas un rētas - ovulācijas pēdas un dzelteno ķermeņu transformācija. Olnīcas svars ir 5-8 g. Olnīcas izmēri ir: garums 2,5-5,5 cm, platums 1,5-3,0 cm, biezums - līdz 2 cm. Olnīcai ir divas brīvas virsmas: mediālā virsma (facies medialis), kas vērsta pret iegurņa dobumu, daļēji pārklāta ar olvadu, un sānu virsma (facies lateralis), kas atrodas blakus iegurņa sānu sienai, līdz nedaudz izteiktai ieplakai - olnīcas bedrei. Šī bedre atrodas leņķī starp ārējiem iegurņa asinsvadiem, kas augšpusē pārklāti ar vēderplēvi, un dzemdes un obturatora artērijām apakšā. Aiz olnīcas atbilstošās puses urīnvads retroperitoneāli iet no augšas uz leju.
Olnīcas virsmas pāriet izliektā brīvā (aizmugurējā) malā (margo liber), priekšpusē - mezentērija malā (margo mesovaricus), kas ar īsu vēderplēves (olnīcas mezentērijas) kroku piestiprināta pie dzemdes platās saites aizmugurējās lapiņas. Uz šīs orgāna priekšējās malas ir rievota ieplaka - olnīcas hilum (hilum ovarii), caur kuru olnīcā iekļūst artērija un nervi, bet iziet vēnas un limfvadi. Olnīcai ir arī divi gali: noapaļotais augšējais cauruļveida gals (extremitas tubaria), kas vērsts pret olvadu, un apakšējais dzemdes gals (extremitas utenna), kas savienots ar dzemdi ar olnīcas saiti (lig. ovarii proprium). Šī saite, apmēram 6 mm biezas apaļas auklas formā, stiepjas no olnīcas dzemdes gala līdz dzemdes sānu stūrim, kas atrodas starp divām dzemdes platās saites lapiņām. Olnīcas saišu aparātā ietilpst arī olnīcas saišu suspensorijs (lig.suspensorium ovarii), kas ir vēderplēves kroka, kas stiepjas no mazā iegurņa sienas līdz olnīcai un kuras iekšpusē atrodas olnīcas asinsvadi un šķiedru šķiedru kūlīši. Olnīcu fiksē īss mezentērijs (mezovārijs), kas ir vēderplēves dubultojums, kas stiepjas no dzemdes platās saites aizmugurējās lapiņas līdz olnīcas mezentērija malai. Pašas olnīcas nav pārklātas ar vēderplēvi. Lielākā olvadu apmale ir piestiprināta pie olnīcas cauruļveida gala. Olnīcas topogrāfija ir atkarīga no dzemdes stāvokļa, tās lieluma (grūtniecības laikā). Olnīcas ir ļoti kustīgi mazā iegurņa dobuma orgāni.
 [
[ Olnīcu asinsvadi un nervi
Olnīcu asinsapgādi nodrošina aa. et vv. ovaricae et uterinae. Abas olnīcu artērijas (aa. ovaricae dextra et sinistra) sākas no aortas priekšējās virsmas tieši zem nieru artērijām; labā artērija parasti sākas no aortas, bet kreisā - no nieru artērijas. Vēršoties uz leju un sāniski gar lielā jostas muskuļa priekšējo virsmu, katra olnīcas artērija šķērso priekšā esošo urīnvadu (atdodot tam zarus), ārējos iegurņa asinsvadus, robežlīniju un nonāk iegurņa dobumā, atrodoties šeit olnīcas suspensorajā saitē. Sekojot mediālā virzienā, olnīcas artērija iet starp dzemdes platās saites lāsītēm zem olvada, atdodot tam zarus, un pēc tam olnīcas mezentērijā; tā nonāk olnīcas hilumā.
Olnīcu artērijas zari plaši anastomozējas ar dzemdes artērijas olnīcu zariem. Venozā izplūde no olnīcām galvenokārt notiek olnīcu venozajā pinumā, kas atrodas olnīcu paugura rajonā. No šejienes asiņu izplūde notiek divos virzienos: caur dzemdes un olnīcu vēnām. Labajai olnīcu vēnai ir vārsti, un tā ieplūst apakšējā dobajā vēnā. Kreisā olnīcu vēna ieplūst kreisajā nieru vēnā, lai gan tai nav vārstuļu.
Limfas attece no olnīcām notiek caur limfvadiem, īpaši bagātīgi orgānu vārtu rajonā, kur izceļas subovāriskais limfātiskais pinums. Pēc tam limfa pa olnīcu limfvadiem nonāk paraaortiskajos limfmezglos.
Olnīcu inervācija
Simpātiskais - ko nodrošina postganglionālās šķiedras no celiakijas (saules), augšējā mezentērija un hipogastriskā pinuma; parasimpātiskais - ko nodrošina viscerālie sakrālie nervi.
Olnīcas struktūra
Olnīcas virsmu klāj vienslāņa dīgļepitēlija slānis. Zem tā atrodas blīvs saistaudu olbaltumvielu apvalks (tunica albuginea). Olnīcas saistaudi veido tās stromu (stroma ovarii), kas bagāta ar elastīgām šķiedrām. Olnīcas viela, tās parenhīma, ir sadalīta ārējā un iekšējā slānī. Iekšējais slānis, kas atrodas olnīcas centrā, tuvāk tās vārtiem, tiek saukts par serdi (medulla ovarii). Šajā slānī, irdenajos saistaudos, atrodas daudzi asinsvadi un limfvadi, kā arī nervi. Olnīcas ārējais slānis - garoza (cortex ovarii) ir blīvāka. Tajā ir daudz saistaudu, kuros atrodas nobriestošie primārie olnīcu folikuli (folliculi ovarici primarii), sekundārie (vezikulārie) folikuli (folliculi ovarici secundarii, s.vesiculosi), kā arī nobrieduši folikuli, Grāfa folikuli (folliculi ovarici maturis), kā arī dzeltenie un atrētiskie ķermeņi.
Katrā folikulā ir sievišķā reproduktīvā olšūna jeb oocīts (oocīts). Olšūna ir līdz 150 µm diametrā, apaļa, satur kodolu, lielu daudzumu citoplazmas, kas papildus šūnu organellām satur olbaltumvielu-lipīdu ieslēgumus (dzeltenumu), glikogēnu, kas nepieciešami olšūnas barošanai. Olšūna parasti izlieto savas barības vielu rezerves 12–24 stundu laikā pēc ovulācijas. Ja apaugļošanās nenotiek, olšūna iet bojā.
Cilvēka olšūnu klāj divas membrānas. Iekšpusē atrodas citolemma, kas ir olšūnas citoplazmas membrāna. Ārpus citolemmas atrodas tā saukto folikulāro šūnu slānis, kas aizsargā olšūnu un veic hormonu veidošanās funkciju – tās izdala estrogēnus.
Dzemdes, olvadu un olnīcu fizioloģisko stāvokli nodrošina piekares, fiksācijas un atbalsta aparāti, kas apvieno vēderplēvi, saites un iegurņa audus. Piekares aparātu attēlo pāru veidojumi, tas ietver dzemdes apaļās un platās saites, olnīcu pareizās saites un piekares saites. Dzemdes platās saites, olnīcu pareizās un piekares saites notur dzemdi vidējā stāvoklī. Apaļās saites velk dzemdes dibenu uz priekšu un nodrošina tās fizioloģisko slīpumu.
Fiksācijas (enkurošanas) aparāts nodrošina dzemdes novietojumu mazā iegurņa centrā un praktiski neļauj tai nobīdīties uz sāniem, uz priekšu un atpakaļ. Taču, tā kā saišu aparāts atiet no dzemdes tās apakšējā daļā, ir iespējama dzemdes noliekšanās dažādos virzienos. Fiksācijas aparāts ietver saites, kas atrodas iegurņa vaļīgajos audos un stiepjas no dzemdes apakšējās daļas līdz iegurņa sānu, priekšējai un aizmugurējai sienai: sakromaģiskās, kardinālās, uterovezikālās un vezikopubiskās saites.
Papildus mezovarijam izšķir šādas olnīcu saites:
- olnīcas suspensorā saite, agrāk apzīmēta kā infundibuloplevic saite. Tā ir vēderplēves kroka ar tai cauri ejošiem asinsvadiem (a. et v. ovarica) un limfvadiem, kā arī olnīcas nerviem, kas izstiepta starp iegurņa sānu sienu, jostas fasciju (kopīgās iegurņa artērijas dalīšanās zonā ārējā un iekšējā) un olnīcas augšējo (olvadu) galu;
- Olnīcas pareizā saite iet starp platās dzemdes saites lapām, tuvāk aizmugurējai lapai, un savieno olnīcas apakšējo galu ar dzemdes sānu malu. Olnīcas pareizā saite ir piestiprināta pie dzemdes starp olvadu sākumu un apaļo saiti, aiz un virs pēdējās. Saites biezumā iet rr. ovarii, kas ir dzemdes artērijas gala zari;
- Apendekālā-olnīcu saite Saite stiepjas no apendiksa mezentērijas līdz labajai olnīcai vai dzemdes platajai saitei vēderplēves krokas veidā. Saite ir nepastāvīga un novērojama 1/2 - 1/3 sieviešu.
Atbalsta aparātu attēlo iegurņa pamatnes muskuļi un fascija, kas sadalīta apakšējā, vidējā un augšējā (iekšējā) slānī.
Visspēcīgākais ir augšējais (iekšējais) muskuļu slānis, ko pārstāv pāru muskulis, kas paceļ anālo atveri. Tas sastāv no muskuļu kūlīšiem, kas izplešas no astes kaula līdz iegurņa kauliem trīs virzienos (pubococcygeus, iliococcygeus un ischiococcygeus). Šo muskuļu slāni sauc arī par iegurņa diafragmu.
Vidējais muskuļu slānis atrodas starp simfizu, kaunuma kaulu un sēžas kaulu. Vidējais muskuļu slānis - uroģenitālā diafragma - aizņem iegurņa izejas priekšējo pusi, caur kuru iziet urīnizvadkanāls un maksts. Priekšējā daļā starp tās loksnēm atrodas muskuļu saišķi, kas veido urīnizvadkanāla ārējo sfinkteru, aizmugurējā daļā - muskuļu saišķi, kas iet šķērsvirzienā - starpenes dziļais šķērsvirziena muskulis.
Iegurņa pamatnes muskuļu apakšējais (ārējais) slānis sastāv no virspusējiem muskuļiem, kuru forma atgādina skaitli 8. Tie ietver bulbokavernozo, išhiokavernozo, ārējo anālo sfinkteru un virspusējo šķērsvirziena starpenes muskuli.
Olnīcu ontoģenēze
Folikulu augšanas un atrēzijas process sākas 20. grūtniecības nedēļā, un līdz dzemdību brīdim meitenes olnīcās paliek līdz 2 miljoniem oocītu. Līdz menstruāciju sākumam to skaits samazinās līdz 300 tūkstošiem. Visā reproduktīvās dzīves periodā nobriest un ovulē ne vairāk kā 500 folikulu. Folikulu sākotnējā augšana nav atkarīga no FSH stimulācijas, ir ierobežota, un atrēzija notiek ātri. Tiek uzskatīts, ka primāro folikulu augšanas un atrēzijas galvenais regulators ir nevis steroīdie hormoni, bet gan lokālie autokrīnie/parakrīnie peptīdi. Tiek uzskatīts, ka folikulu augšanas un atrēzijas procesu nepārtrauc nekādi fizioloģiski procesi. Šis process turpinās visos vecumos, ieskaitot intrauterīnu periodu un menopauzi, un to pārtrauc grūtniecība, ovulācija un anovulācija. Mehānisms, kas izraisa folikulu augšanu un to skaitu katrā konkrētajā ciklā, joprojām nav skaidrs.
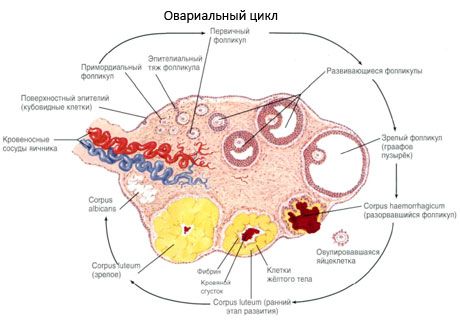
Attīstības laikā folikuls iziet vairākas attīstības stadijas. Primordiālās dīgļšūnas rodas no dzeltenuma maisiņa endodermas, alantoisa, un 5.–6. grūtniecības nedēļā migrē uz embrija dzimumorgānu apvidu. Ātras mitotiskas dalīšanās rezultātā, kas turpinās no 6.–8. grūtniecības nedēļas līdz 16.–20. grūtniecības nedēļai, embrija olnīcās veidojas līdz pat 6–7 miljoniem olšūnu, ko ieskauj plāns granulāro šūnu slānis.
Preantrāls folikuls - olšūnu ieskauj membrāna (Zona pellucida). Granulozes šūnas, kas ieskauj olšūnu, sāk vairoties, to augšana ir atkarīga no gonadotropīniem un korelē ar estrogēnu līmeni. Granulozes šūnas ir FSH mērķis. Preantrālā folikula stadijā granulozes šūnas spēj sintezēt trīs steroīdu klases: galvenokārt inducē aromatāzes, galvenā enzīma, kas pārvērš androgēnus par estradiolu, aktivitāti. Tiek uzskatīts, ka estradiols spēj palielināt savu receptoru skaitu, radot tiešu mitogēnu iedarbību uz granulozes šūnām neatkarīgi no FSH. Tas tiek uzskatīts par parakrīnu faktoru, kas pastiprina FSH iedarbību, tostarp aromatizācijas procesu aktivizēšanu.
FSH receptori parādās uz granulāro šūnu membrānām, tiklīdz sākas folikulu augšana. FSH samazināšanās vai palielināšanās izraisa tā receptoru skaita izmaiņas. Šo FSH darbību modulē augšanas faktori. FSH darbojas caur G-proteīnu, adenilātciklāzes sistēmu, lai gan steroīdu veidošanos folikulā galvenokārt regulē FSH, šajā procesā ir iesaistīti daudzi faktori: jonu kanāli, tirozīnkināzes receptori, otro kurjeru fosfolipāzes sistēma.
Androgēnu loma agrīnā folikulu attīstībā ir sarežģīta. Granulozām šūnām ir androgēnu receptori. Tie ir ne tikai substrāts FSH izraisītai aromatizācijai par estrogēniem, bet arī var pastiprināt aromatizāciju zemās koncentrācijās. Kad androgēnu līmenis palielinās, preantrālās granulozās šūnas priekšroku dod nevis aromatizācijas ceļam uz estrogēniem, bet gan vienkāršākam androgēnu ceļam caur 5α-reduktāzi, lai pārvērstu par androgēnu, ko nevar pārvērst par estrogēnu, tādējādi kavējot aromatāzes aktivitāti. Šis process arī kavē FSH un LH receptoru veidošanos, tādējādi apturot folikulu attīstību.
Aromatizācijas procesā folikuls ar augstu androgēnu līmeni piedzīvo atrēzijas procesus. Folikula augšana un attīstība ir atkarīga no tā spējas pārvērst androgēnus par estrogēniem.
FSH klātbūtnē folikulu šķidruma dominējošā viela būs estrogēni. FSH neesamības gadījumā - androgēni. LH parasti folikulu šķidrumā nav līdz cikla vidum. Tiklīdz LH līmenis paaugstinās, granulāro šūnu mitotiskā aktivitāte samazinās, parādās deģeneratīvas izmaiņas un paaugstinās androgēnu līmenis folikulā. Steroīdu līmenis folikulu šķidrumā ir augstāks nekā plazmā un atspoguļo olnīcu šūnu: granulāro un teka šūnu funkcionālo aktivitāti. Ja vienīgais FSH mērķis ir granulārās šūnas, tad LH ir daudz mērķu - teka šūnas, stromas un luteīna šūnas un granulārās šūnas. Gan granulārajām, gan teka šūnām piemīt spēja steroīdu sintēzei, bet granulārajās šūnās dominē aromatāzes aktivitāte.
Reaģējot uz LH, teka šūnas ražo androgēnus, kurus pēc tam granulāras šūnas FSH inducētas aromatizācijas rezultātā pārvērš estrogēnos.
Folikulam augot, teka šūnas sāk ekspresēt LH receptoru, P450 sec un 3beta-hidroksisteroīdu dehidrogenāzes gēnus, insulīnam līdzīgais augšanas faktors (IGF-1) sinerģiski ar LH palielina gēnu ekspresiju, bet nestimulē steroīdu veidošanos.
Olnīcu steroīdu veidošanās vienmēr ir atkarīga no LH. Folikulam augot, teka šūnas ekspresē enzīmu P450c17, kas no holesterīna veido androgēnus. Granulosa šūnām šī enzīma nav, un tās ir atkarīgas no teka šūnām, lai tās no androgēniem ražotu estrogēnus. Atšķirībā no steroīdu veidošanās, folikuloģenēze ir atkarīga no FSH. Folikulam augot un estrogēna līmenim palielinoties, tiek aktivizēts atgriezeniskās saites mehānisms - FSH veidošanās tiek kavēta, kas savukārt noved pie folikula aromatāzes aktivitātes samazināšanās un galu galā pie folikulu atrēzijas, izmantojot apoptozi (ieprogrammētu šūnu nāvi).
Estrogēnu un FSH atgriezeniskās saites mehānisms kavē jau sākušos folikulu attīstību, bet ne dominējošā folikula attīstību. Dominējošais folikuls satur vairāk FSH receptoru, kas atbalsta granulāro šūnu proliferāciju un androgēnu aromatizāciju par estrogēniem. Turklāt parakrīnie un autokrīnie ceļi darbojas kā svarīgs antrālo folikulu attīstības koordinators.
Autokrīnais/parakrīnais regulators sastāv no peptīdiem (inhibīna, aktivīna, folistatīna), ko sintezē granulāras šūnas, reaģējot uz FSH, un kas nonāk folikulu šķidrumā. Inhibīns samazina FSH sekrēciju; aktivīns stimulē FSH izdalīšanos no hipofīzes un pastiprina FSH darbību olnīcā; folistatīns nomāc FSH aktivitāti, iespējams, saistoties ar aktivīnu. Pēc ovulācijas un dzeltenā ķermeņa attīstības inhibīnu kontrolē LH.
Olnīcu šūnu augšanu un diferenciāciju ietekmē insulīnam līdzīgie augšanas faktori (IGE). IGF-1 iedarbojas uz granulārajām šūnām, izraisot cikliskā adenozīna monofosfāta (cAMP), progesterona, oksitocīna, proteoglikāna un inhibīna līmeņa paaugstināšanos.
IGF-1 iedarbojas uz teka šūnām, izraisot palielinātu androgēnu veidošanos. Teka šūnas savukārt ražo audzēja nekrozes faktoru (TNF) un epidermas augšanas faktoru (EGF), kurus arī regulē FSH.
EGF stimulē granulāro šūnu proliferāciju. IGF-2 ir galvenais augšanas faktors folikulu šķidrumā, un tajā ir atrodami arī IGF-1, TNF-α, TNF-3 un EGF.
Šķiet, ka ovulācijas traucējumu un policistisko olnīcu attīstības gadījumos nozīme ir olnīcu funkcijas parakrīnās un/vai autokrīnās regulācijas traucējumiem.
Antrālajam folikulam augot, folikulu šķidrumā palielinās estrogēna saturs. To pieauguma kulminācijā uz granulārajām šūnām parādās LH receptori, notiek granulāras šūnu luteinizācija un palielinās progesterona ražošana. Tādējādi preovulācijas periodā estrogēna ražošanas palielināšanās izraisa LH receptoru parādīšanos, LH savukārt izraisa granulāras šūnu luteinizāciju un progesterona ražošanu. Progesterona līmeņa paaugstināšanās samazina estrogēna līmeni, kas acīmredzot izraisa otro FSH maksimumu cikla vidū.
Tiek uzskatīts, ka ovulācija notiek 10–12 stundas pēc LH maksimuma un 24–36 stundas pēc estradiola maksimuma. Tiek uzskatīts, ka LH stimulē olšūnas dalīšanos, granulāro šūnu luteinizāciju un progesterona un prostaglandīna sintēzi folikulā.
Progesterons pastiprina proteolītisko enzīmu aktivitāti, kas kopā ar prostaglandīnu piedalās folikula sieniņas plīsumā. Progesterona izraisītais FSH maksimums ļauj oocītam iziet no folikula, pārvēršot plazminogēnu par proteolītisko enzīmu plazmīnu, un nodrošina pietiekamu LH receptoru skaitu luteālās fāzes normālai attīstībai.
3 dienu laikā pēc ovulācijas granulāras šūnas palielinās izmēros, un tajās parādās raksturīgas vakuolas, kas pildītas ar pigmentu luteīnu. Teka-luteālās šūnas diferencējas no tekas un stromas un kļūst par dzeltenā ķermeņa daļu. Kapilāri, kas iekļūst dzeltenajā ķermenī, angioģenēzes faktoru ietekmē attīstās ļoti ātri, un, uzlabojoties vaskularizācijai, palielinās progesterona un estrogēnu ražošana. Steroīdu sintēzes aktivitāti un dzeltenā ķermeņa dzīves ilgumu nosaka LH līmenis. Dzeltenais ķermenis nav homogēns šūnu veidojums. Papildus 2 dzeltenā ķermeņa veidiem tas satur endotēlija šūnas, makrofāgus, fibroblastus utt. Lielas dzeltenā ķermeņa šūnas ražo peptīdus (relaksīnu, oksitocīnu) un ir aktīvākas steroīdu ģenēzē ar lielāku aromatāzes aktivitāti un lielāku progesterona sintēzi nekā mazās šūnas.
Progesterona maksimums ir novērojams 8. dienā pēc LH maksimuma sasniegšanas. Jāatzīmē, ka progesterons un estradiols luteālajā fāzē tiek izdalīti epizodiski korelācijā ar LH pulsējošo izdalīšanos. Veidojoties dzeltenajam ķermenim, inhibīna ražošanas kontrole pāriet no FSH uz LH. Inhibīna līmenis palielinās līdz ar estradiola līmeņa paaugstināšanos pirms LH maksimuma sasniegšanas un turpina pieaugt pēc LH maksimuma sasniegšanas, lai gan estrogēna līmenis samazinās. Lai gan inhibīnu un estradiolu izdala granulārās šūnas, to regulēšanu veic dažādi ceļi. Inhibīna līmeņa pazemināšanās luteālās fāzes beigās veicina FSH līmeņa paaugstināšanos nākamajā ciklā.
Dzeltenais ķermenis samazinās ļoti ātri - 9.-11. dienā pēc ovulācijas.
Deģenerācijas mehānisms nav skaidrs un nav saistīts ar estrogēnu luteolītisko lomu vai ar receptoru saistītu mehānismu, kā novērots endometrijā. Pastāv vēl viens skaidrojums dzeltenā ķermeņa ražoto estrogēnu lomai. Ir zināms, ka estrogēni ir nepieciešami progesterona receptoru sintēzei endometrijā. Luteālās fāzes estrogēni, iespējams, ir nepieciešami ar progesteronu saistītām izmaiņām endometrijā pēc ovulācijas. Nepietiekama progesterona receptoru attīstība nepietiekama estrogēna līmeņa rezultātā, iespējams, ir papildu mehānisms neauglībai un agrīniem abortiem, kas ir vēl viena luteālās fāzes deficīta forma. Tiek uzskatīts, ka dzeltenā ķermeņa dzīves ilgums tiek noteikts ovulācijas laikā. Un tas noteikti regresēs, ja grūtniecības dēļ to neatbalsta cilvēka horiona gonadotropīns. Tādējādi dzeltenā ķermeņa regresija noved pie estradiola, progesterona un inhibīna līmeņa pazemināšanās. Inhibīna līmeņa pazemināšanās novērš tā nomācošo iedarbību uz FSH; Estradiola un progesterona līmeņa pazemināšanās ļauj ļoti ātri atjaunot GnRH sekrēciju un novērst atgriezeniskās saites mehānismu no hipofīzes. Inhibīna un estradiola līmeņa pazemināšanās kopā ar GnRH līmeņa paaugstināšanos izraisa FSH veidošanos, nevis LH. FSH līmeņa paaugstināšanās izraisa folikulu augšanu, kam seko dominējošā folikula atlase, un, ja grūtniecība neiestājas, sākas jauns cikls. Steroīdu hormoniem ir vadoša loma reproduktīvajā bioloģijā un vispārējā fizioloģijā. Tie nosaka cilvēka fenotipu, ietekmē sirds un asinsvadu sistēmu, kaulu metabolismu, ādu, vispārējo labsajūtu un tiem ir galvenā loma grūtniecības laikā. Steroīdu hormonu darbība atspoguļo intracelulāros un ģenētiskos mehānismus, kas nepieciešami, lai pārraidītu ekstracelulāru signālu uz šūnas kodolu, lai izraisītu fizioloģisku reakciju.
Estrogēni difūzā veidā iekļūst šūnu membrānā un saistās ar receptoriem, kas atrodas šūnas kodolā. Pēc tam receptoru-steroīdu komplekss saistās ar DNS. Mērķa šūnās šī mijiedarbība noved pie gēnu ekspresijas, olbaltumvielu sintēzes un specifiskām šūnu un audu funkcijām.
Использованная литература